 Figure 1. Left to right. Phil Sheridan, Bill Scholl, John Hummer, Edwin Bridges and Steve
Figure 1. Left to right. Phil Sheridan, Bill Scholl, John Hummer, Edwin Bridges and Steve Orzell, in the historic meeting at Sandy Creek near Wewahitchka, Florida, May 12, 1987.
Print Page | Close Page |
Home
Carnivorous Plant Newsletter 20: 102-110
by Philip M. Sheridan
Meadowview Biological Research Station
8390 Fredericksburg Turnpike
Woodford, VA 22580
meadowview@pitcherplant.org
Abstract
The distribution, variation and habitats of Sarracenia alata Wood in the West Gulf Coastal Plain are discussed. The relationships and possible origin of this species are also discussed.
Introduction
Visits to Sarracenia alata Wood sites in Mississippi and Alabama from 1984-1987, and growing this species since 1974, convinced me of the need to examine its range in Texas and western Louisiana for its full variation. This investigation was necessary since there is very little published information on S. alata in the westem part of its range (MacRoberts and MacRoberts, 1988). It has been an overlooked pitcher plant for scientific study and its exact placement in Sarracenia phylogeny is not understood.
While botanizing in May of 1987 near Wewahitchka in western Florida with fellow naturalists Bill Scholl and John Hummer, I had the good fortune to meet two botanists from Texas, Edwin Bridges and Steve Orzell (Figure 1). Messrs. Bridges and Orzell were studying wetlands in the West Gulf Coastal Plain and knew of many western S. alata sites in Texas and western Louisiana. This encounter resulted in an invitation to join them on a field survey of these westernmost bogs from June 7-12,1988.
Additional visits to Texas and western Louisiana were made from June 19-20, 1989 with Mr.John Hummer. Twenty-two sites in nine counties and five parishes were investigated from 1988-1989 out of a total of approximately 85 extant stations for S. alata in the West Gulf Coastal Plain (Bridges and Orzell, pers. comm. 1989). Unreported information about the western stations were discovered on these trips.
Figure 1. Left to right. Phil Sheridan, Bill Scholl, John Hummer, Edwin Bridges and Steve
Distribution
Texas accounts for sixteen of the thirty-six county records in four states for S. alata. This species has also been found in seven Louisiana parishes, ten Mississippi and three Alabama counties. With a total of twenty counties and parishes (more than half the range) in Texas and western Louisiana, it is truly the West Gulf Coast Pitcher Plant.
The range of S. alata is bicentric with a gap of one hundred and forty miles in central Louisiana (Figure 2). Unsuitable soils and alluvial swamp do not allow its survival and colonization in central Louisiana which also does not have any seepage bog or wetland savannah habitats. The range is limited in the east by the Alabama/ Tombigbee Rivers (with the exception of Baldwin County, Alabama), on the south by the Gulf of Mexico, and by lower annual rainfall to the west. The northern range for S. alata appears to be limited by the lack of suitable habitat, such as pine savannas and acid hillside seepage bogs, and competition from other plants.
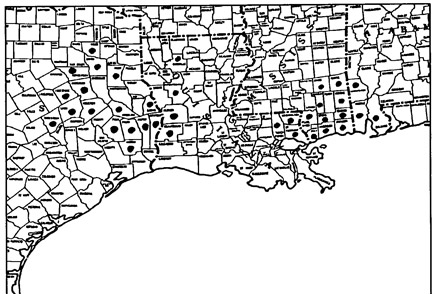 Figure 2. Distribution of S. alata
Figure 2. Distribution of S. alata
Some older records for S. alata in the West Gulf Coastal Plain are questionable. Calcasieu Parish in Louisiana included present-day Beauregard Parish in the 1800s and it is likely that an old collection there was actually made in Beauregard Parish. Collections from San Augustine County, Texas may be mislabeled since their location data leads to sites in Angelina and Jasper counties, Texas. No extant natural populations are known from Hardin, Rush, Smith, Houston or San Augustine counties in Texas. It is also very rare in Bienville Parish, Louisiana and Sabine and Robertson counties, Texas, known only from one station per county. It is also very rare in Wood County, Texas with only two sites (Bridges and Orzell, pers. comm. 1989) and is locally abundant in Angelina, Jasper and Newton Counties, Texas and Vernon and Beauregard Parishes, Louisiana.
Volume 20 December 1991 (103)
Variation and Habitat
Sarracenia alata bogs in the western-part of the range are remarkable because they are located in a region with no native pine species. Most notably absent is longleaf pine, Pinus palustris. Annual rainfall is insufficient to support pine trees, but allows a variety of oaks such as Quercus stellata Wang., Q. incana Bartram and Q. marilandica Muenchh. to flourish. The most western bogs, especially in Wood, Robertson and Leon counties, are kept at constant moisture levels by spring water and have been classified as deep seepage bogs (Bridges and Orzell, 1989). In contrast, many East Gulf Coastal Plain wetland pine savanna habitats receive much of their water from rainfall.
In Robertson, Henderson, Wood and Anderson counties, in east central Texas, pitchers are green to greenish yellow in color with moderate purple veining. The pitchers have noticeable expansion of the upper pitcher tube and arching hoods. Some of the larger specimens measure 30 inches tall with hood measurements of 2 inches in width and 3 inches in length. The Robertson County, Texas site contains several thousand plants, is the only site known in the county, the most western in the species range and less than 100 miles from Austin, Texas! This site, like many S. alata bogs in the western part of the range, is being invaded by woody vegetation and needs prescribed burning to restore herbaceous plant vigor.
A site examined in Leon County, in central Texas, contained a number of plants with deep purple pigmentation of the veins, lip and orifice of the pitcher (Figure 3). Some leaves were almost completely reddish-purple. This extreme coloration is attributed to the open habitat and age of the leaves. Fresh leaves, while having heavy pigmentation, were not completely reddish-purple. This heavy pigmentation appears to be an infrequent feature in the western stations of S. alata, although commonly found at sites in Mississippi and Alabama. The dominant color form in the most western of the S. alata bogs is a green pitcher with moderate purple venation inside the trumpet (Figure 4).
Sarracenia alata is found naturally in wetland longleaf pine savanna at only one site in Beauregard Parish, Louisiana although it has been introduced in this habitat in the Big Thicket National Preserve in Tyler and Hardin counties, Texas (Bridges and Orzell, 1989). Wetland longleaf pine savannas have a seasonally high water table due to low topographic position or an impermeable soil layer. The soil has a silt or clay surface (Bridges and Orzell, 1989). However, it is commonly found in clay-based savannas in southern Mississippi (Norquist, 1984) and southeastern Louisiana (Bridges and Orzell, 1989). The wetland savannah in Beauregard Parish, Louisiana contains plants which are unusual in their undulate hood, shape of the pitcher mouth and the olive or greenish yellow pitcher color with reddish color on the top of the hood. In many respects they are similar to S. alabamensis ssp. wheryyii. These differences are noteworthy considering this is the only native site for S. alata in wetland longleaf pine savannas of Texas and western Louisiana. Sarracenia alata is occasionally found in semi-evergreen broadleaf acid seep forests in Texas and western Louisiana. These sites "occupy narrow areas along upper reaches of streams in sandy areas of the Longleaf Pine Savannas Section. This type also occurs, but less frequently, in the counties immediately north of this section," (Bridges and Orzell, 1989). It has a greater dominance and richness of evergreens in the shrub and canopy layers. Sarracenia alata, as well as other hillside seepage bog species, are found in few numbers on the fringes of this type of habitat and maybe relics of a time when these forests were maintained in an open state by natural fires. The most northern sites for the species in Bienville and Natchitoches Parish, Louisiana are semi-evergreen broadleaf acid seep forests.
Bienville Parish, Louisiana is two parishes south of Arkansas and represents the most northern limit for pitcher plant bogs in Louisiana. A dense canopy of Magnolia virginiana L., Nyssa sylvatica var biflora (Walter) Sargent, Acer rubrum
Volume 20 December 1991 (104)
L. and Liquidambar styraciflua L., as well as competition from shrubs, allowed limited sunlight to reach the pitcher plants in the one site examined. As a result of insufficient sunlight, the plants were in a depauperate condition. They appeared to be green pitchers with moderate purple veining.
A former semi-evergreen broadleaf acid seep forest examined in Natchitoches Parish, Louisiana which has now been converted to pasture land contains no more than fifteen clumps of S. alata. Some of these clumps, however, measured three feet by six feet with individual leaves up to thirty-five inches tall! The flower stalks were noteworthy because of their uniform short length of only nine inches when compared to most pitcher heights of almost three feet. Associates were Nyssa sylvatica var biflora, Solidago patula Muhl. ex Willid., Saururus cernuus L. and Rhynchospora rariflora (Michaux) Ell.
Sarracenia alata occurs in about 68% of the hillside seepage bogs in the West Gulf coastal plain (Bridges and Orzell, 1989). These bogs are usually "found on short, steep slopes (10 to 30 percent), generally near midslope of the headwaters of small ravines but occasionally on lower slopes at headwaters of small streams …" The substrate is usually a loamy sand or sandy loam, sometimes developing into a sandy peat or shallow mucky peat," (Bridges and Orzell, 1989). The flora is mainly graminoid with wetland forbs and occasional broadleaf evergreen shrubs and lianas. These seepage bogs are "restricted to the Longleaf Pine Savannas Section of the West Gulf Coastal Plain, from Natchitoches Parish, Louisiana, west to Polk County, Texas, in areas of relatively rolling terrain. Individual bogs are generally less than 2 ha. in size, with many less than 0.4 ha. but often several occur within a site. In Louisiana representative examples occur in the Kisatchie National Forest and in western Beauregard Parish. In Texas, bogs occur most frequently in northern Newton and Jasper counties, with the largest concentration of high quality bogs at the Willis/Catahoula Formation contact in the southern part of the Angelina National Forest." (Bridges and Orzell, 1989).
Soil and temperature were analyzed in one hillside seepage bog in Jasper County, Texas on June 19, 1989 resulted in a pH of 5.5., nitrogen and phosphorus at approximately 4% with negligible amounts of potash. Air temperature at midday was 90 degrees F. with a soil temperature two inches underground in full sun of 89 degrees F. Soil temperature in the shade two inches underground was 84 degrees F.
Sarracenia alata found in hillside seepage bogs are golden yellow in color (Figure 5) with widely expanded upper pitchers. Moderate purple venation is found inside the pitcher. As the pitchers age a tan color may develop on the hood and exterior pitcher. Robust specimens can measure 33 inches tall with hood lengths and widths of 3.5 inches.
Flower color observed on cultivated specimens of S. alata from hillside seepage bogs is cream yellow. Flowers seem to have a clean or fresh odor. Eleven of my friends and co-workers were surveyed for their opinion of the odor of the flowers. A flower was placed in a glass jar and exposed to moderate sunlight. Five of these people detected a clean or fresh spring meadow scent. One individual noted the classic domestic cat urine smell noted in Schnell (1978), while others reported odors of turpentine, "Opium" perfume, fruit, mango leaves or a bitter smell! It is interesting to note the variety of odors reported. I could detect a turpentine odor but could equate this with the spring meadow or fresh odor detected by others. I have detected a reduced domestic cat urine odor associated with a fresh scent. Thus the flower odor of S. alata would seem to be unique, although possibly possessing similarities to S. flava.
The most exciting feature is the presence of areoles or light patches on S. alata in hillside seepage bogs. Plants with areoles were found in the counties of Angelina, Newton and Jasper, Texas and in Natchitoches and Beauregard Parishes, Louisiana.
Volume 20 December 1991 (105)
 Figure 3. Heavily pigmented forms of S. alatain Leon
Figure 3. Heavily pigmented forms of S. alatain Leon
 Figure 4. Typical form of S. alata in the western part of its range. June 7, 1988.
Figure 4. Typical form of S. alata in the western part of its range. June 7, 1988.
 Figure 5. Golden Yellow S. alata in hillside seepage bog of Newton County, Texas. Note Asclepias rubra L., Calopogon tuberosus (L) Bsp., Rudbekia scabrifolia L.E. Brown and Eupatorium rotundiolium L. in flower. June 10, 1988.
Figure 5. Golden Yellow S. alata in hillside seepage bog of Newton County, Texas. Note Asclepias rubra L., Calopogon tuberosus (L) Bsp., Rudbekia scabrifolia L.E. Brown and Eupatorium rotundiolium L. in flower. June 10, 1988.
 Figure 6. Comparison of S. alata (Number 339 Jasper County, Texas; Number 340 Beauregard Parish, Louisiana) and S. alabamensis ssp. alabamensis (Number 341 Chilton County, Alabama). Specimens deposited at Longwood College, Farmville, Virginia (FARM).
Figure 6. Comparison of S. alata (Number 339 Jasper County, Texas; Number 340 Beauregard Parish, Louisiana) and S. alabamensis ssp. alabamensis (Number 341 Chilton County, Alabama). Specimens deposited at Longwood College, Farmville, Virginia (FARM).
Volume 20 December 1991 (106)
Areoles are commonly known in S. Ieucophylla, S. psittacina and S.minor and have been noticed by Case and Case (1976) in S.alabamensis ssp. alabamensis and in S. rubra ssp. Jonesii and S. rubra ssp. rubra by Schnell (1977,1990). The Cases (1974) notes that the areoles in S. alabamensis ssp. alabamensis are " . . . the consequence of the extreme tissue expansion characteristic of the summer pitchers. They are not structurally quite the same as in the truly areolate species." Areoles on S.alata in Texas and western Louisiana are similar to those in S.alabamensis ssp. alabamensis, although not confined to summer leaves but produced on leaves throughout the season. A comparative picture of pressed fall leaves of S. alata and S. alabamensis ssp. alabamensis from cultivated material demonstrates the similarity of the areoles in both species (Figure 6).
Hybridization often occurs between S. alata, S. psittacina and S.Ieucophylla in Alabama and Mississippi where the pitcher plant species ranges overlap. One of the results of this overlap in range is introgressed S. alata with areoles. Whether any of these areolate plants are pure genetic forms and not introgressed hybrids must await further analysis. Introgression, however, appears unlikely to be a factor in the presence of areoles in the Texas and western Louisiana range of S.alata since McDaniel (1966) noted that particular components found in introgressed S.alata were not found in areas where hybridization is improbable. Hybridization between S.alata and any other sarracenia is highly improbable in Texas and western Louisiana since no other pitcher plant now naturally grows in these areas. Thus it appears the presence of areoles in Texas and western Louisiana plants is an inherent feature of the species and not the result of recent introgression with areolate species of sarracenia. Ancient introgression between S.alata and areolate sarracenia cannot be eliminated as a factor in present-day S. alata without chemical, paleobotanical and phytogeographic evidence.
It should be emphasized that the occurrence of areoles in plants of Texas and western Louisiana S. alata stations is a feature which may be enhanced by environmental conditions. Burning of a S.alata bog in Beauregard Parish in 1989 demonstrated this point. A site for S.alata in Beauregard Parish, Louisiana was visited in both June of 1988 and 1989. Although areolate pitchers were sought in 1988, none were seen. Many pitchers were damaged by an Exyra larvae infestation which could have obscured the presence of windows. Areoles were observed in great numbers of plants in June 1989 several months after a fire. Apparently the improved growing conditions provided by fire and suppression of Exyra larvae aided the production of areoles. It should be noted that this bog in Beauregard Parish, Louisiana possess a large, naturally open area. This area in June of 1988 was not noted to contain any obvious areolate plants but did contain them in 1989. The effect of fire, even on an open area, may have been enough to produce areoles. Conceivably the suppression of Exyra larvae by fire played a role in reducing insect damage to pitchers and enhancing areole production.
Pipeline hillside seepage bogs in Jasper County, Texas also illustrate environmental conditions enhancing areole production. Areoles were observed in a Jasper County, Texas pipeline hillside seepage bog in both 1988 and 1989. The site is maintained in a very open condition by annual mowing of the pipeline right-of-way. These annual mowing provide open habitat and allow dense stands of S. alata to develop. Fire would naturally maintain the hillside seepage bog in an open condition, but in this instance the pipeline has substituted as a disturbance factor. Mechanical clearing by bushhogs simulates the effect of fire by inhibiting the growth of woody plants. The suppression of woody plants allows herbaceous types to flourish. Thus, disturbance of hillside seepage bogs may enhance areole production in S. alata by providing open growing conditions.
Volume 20 December 1991 (107)
The occurrence of areoles may be a recent evolutionary phenomenon caused by natural selection. While filming a trapped bumblebee in a S. alata pitcher in a seepage bog in Newton County, Texas in 1988, I noticed that the insect attempted to escape through the sides of the trumpet. Apparently the light-blocking hood of the pitcher confused the insect in how to escape through the open pitcher mouth. Light streaming in through the sides of the leaf caused the bumblebee to fly up and down in the pitcher as if the insect were trying to escape through the translucent pitcher walls. After many attempts to escape, the insect finally collapsed in exhaustion in the recesses of the pitcher. Areoles could further confuse the insect by allowing more light to enter the pitcher. Conceivably, more insects would then be caught. Continued large prey captures would result in more vigorous plants with more seeds and thus a greater likelihood that areolate S. alata will continue to survive, flourish and spread through the process of natural selection.
Relationships within Sarracenia
Several authors have noticed a strong resemblance between S. alata and the S. rubra complex. (Case & Case, 1976; McDaniel, 1966: Schnell 1976, 1978). Other authors (MacFarlane, 1908; Wherry, 1935; Bell, 1949) have aligned S. alata with the S. flava/S. oreophila group. On the S. flava/S. oreophila group Schnell and Krider (1976) state, "Our impression is that this interpretation of relationship was based on the factors of erect pitcher habitus, similarity in pitcher background color (yellow-green to yellow), yellow petals, a weaker though still striking flava-like feline or musty odor, and the tendency for S. alata to grow in similar habitat westward from where the S. flava range ends." "However," they continue, "while these arguments may be superficially compelling—a careful morphological examination in perspective will confirm the closer relationship of S. alata to the S. rubra group, since petal shape, lid shape, and degree of reflexion as well as details of the lid column all more closely resemble the S. rubra group," (Schnell &: Krider, 1976). I completely agree with these observations, and would add that S. alata lacks phyllodia, the flowers do not tilt upward as in S. flava after anthesis and the sepals recurve when the flower matures. These features separate S. alata from the S. flava /S. oreophila group and place it with S. rubra.
The Origin of S. alata
McDaniel (1966) suggests that S. alata arose as a result of hybridization between S. rubra and S. flava or their ancestors. This can be supported by the characteristics that S. alata possesses from both putative parents. Sarracenia alata has the flava-like characteristics of yellowish color, large yellow flower, similar flower odor and early flowering time. It also has the rubra-like characteristics of absence of phyllodia, petal shape, recurvature of sepals, lid shape, degree of reflexion of the hood and details of the lid column. Furthermore, petal shape and general size seems to be intermediate between the two putative parent species. Especially significant to me is the slight feline odor, recurved sepals and petals of the flowers of S. alata. The S. rubra complex possesses flowers which are sweet (Schnell, 1978). The fact that the feline flower odor of the S. flava/S. oreophila group occurs in a pitcher plant with sepals and petals similar to the S. rubra complex is highly suggestive of a hybrid origin for S.alata.
The hybrid origin of S. alata is a fascinating idea yet still needs more scientific data such as statistical analysis of intermediate characteristics, fossilized pollen from area of origin, phytogeographic studies, DNA probe, polymerase chain reaction studies and hybrid experiments to be accepted. If S. alata arose as a hybrid between S.flava/S. Oreophila and the S. rubra complex one would think the hybrid would occasionally still occur in the wild and that does not seem to be the case. I have yet to see a specimen of such a hybrid in the wild or a herbarium. The hybrid, however, may have, arisen from the ancestors of the present species under environmental conditions favorable to hybridization, survival and spread of the new species. Hybrid experiments are now underway between the S. rubra complex and S. flava/S.oreophila by Bill Scholl and me to explore the possible hybrid origin of S.alata further and will be reported in a future paper.
Volume 20 December 1991 (108)
Case and Case (1976) state, "Many pitchers of S. jonesii cannot be distinguished from similarly sized pitchers of S. alata (Wood) Wood. No one seriously considers that S. jonesii belongs to that species." Fred Case, in personal conversations, has since expanded on this original quote to the point that S. alata may indeed be a large, yellow flowered S. jonesii! It is noteworthy that Schnell and Krider ( 1976) found that S. alata was more closely related phenetically to S. jonesii than to any other member of the S. rubra complex. Case and Case (1976) propose a migration route for S. jonesii along the Flint and Coosa Rivers which would have brought it into contact with S. oreophila (or its ancestor) in western Georgia and central Alabama. Recently discovered sites in southwest North Carolina also place S. jonesii very near to S. oreophila (Govus, 1987). Such a contact could have resulted in hybridization and may explain the close phenetic relationship between S. jonesii and S.alata.
It has also been proposed that Sarracenia alata isactually a western S. alabamensis ssp. alabamensis that has evolved a larger yellow flower. Areoles, golden yellow color and general structural features of S. alata could lend support to a relationship with S. alabamensis ssp. alabamensis.
Finally, it is possible that S. alata is simply exhibiting convergent evolution with S. flava/S. oreophila and the S.rubra complex. Natural selection could have resulted in useful features of other pitcher plant species occurring in S. alata. The presence of areoles may be an example of such an event actually taking place.
The question remains though, what is the origin of the West Gulf coast pitcher plant? Is it a western S. alabamensis ssp. alabamensis, a large yellow flowered S. jonesii, a hybrid between ancestral stock of S. rubra and S. flava/S. oreophila or a more distantly related species? This question warrants further investigation.
Conclusion
Sarracenia alata is distributed in the West Gulf coastal plain with very local sites at the extremes of the range. The major color form is green to golden yellow with moderate purple veining inside the trumpet. Heavily pigmented forms are infrequent. Flower color is cream/yellow, with the flowers having an odor similar to S. flava but are unique in having a spring meadow or fresh scent. It mainly occupies hillside seepage bogs and deep muck seepage bogs but also grows in semi-evergreen acid seep forests and is rarely found in wetland longleaf pine savannas. Areoles are found in S. alata growing in hillside seepage bogs. It is closely related to the S. rubra complex and may have descended from a hybrid between the S. flava/S. oreophila and the S.rubra complex or their ancestors.
Note: VHS videotapes are available from the author for $29.95, plus postage, on the above subject.
Volume 20 December 1991 (109)
Acknowledgements
I would especially like to thank Edwin Bridges and Steve Orzell for their invaluable assistance. Additional thanks are due to Dr. Alton Harvill, Don Schnell, John Hummer, Ruth Curlee and Bill Scholl for their comments, ideas and review.
Bibliography
Bell, C.R. 1949 A cytotaxonomic study of the Sarraceniaceae of North America. J. Elisha Mitch. Soc. 65:137-66.
Bridges, Edwin and Orzell, Steve. Personal communication 2/24/89.
Bridges, Edwin and Orzell, Longleaf Pine Communities of the West Gulf Coastal Plain. Natural Areas Journal.9(4).:246-263.
Case, F.W., and R.B. Case.1974. Sarracenia alabamensis, a newly recognized species from central Alabama. Rhodora. 76:650-665.
_____1976. The Sarracenia rubra complex. Rhodora. 78:270-325.
Govus, Thomas E.1987. The occurrence of Sarracenia oreophila (Kearney) Wherry in the blue ridge province of southwestern North Carolina. Castenea 52:310-311.
McDaniel, S.T. 1966. A taxonomic revision of Sarracenia (Sarraceniaceae). Unpubl. PhD Diss., Florida State Univ. (Available from Univ. Microfilms, Ann Arbor, Mich.)
MacFarlane, J.M. 1908. An Engler, Das Pflanzenreich 4:pt. 110.
MacRoberts, B.R. and M.H. MacRoberts. 1988. Floristic composition of two west Louisiana pitcher plant bogs. Phytologia 65:184-190.
Norquist, H.C. 1984. A comparative study of the soils and vegetation of savannas in Mississippi. M.S. Thesis. Mississippi State University, Mississippi State, Miss. IlOp.
Schnell, D.E. and Krider, Daniel W. 1976. Cluster Analysis of the Genus Sarracenia L. in the Southeastern United States. Castanea 41:165-176.
Schnell, D.E. 1977. Infraspecific variation in Sarracenia rubra Walt: Some observations. Castanea 42:149-170.
Schnell, D.E.1978. Sarracenia L. Petal Extract Chromatography. Castanea 43:107115.
Schnell, D.E.1978. Systematic Flower Studies of Sarracenia L. Castanea 43:211-220.
Schnell, D.E. 1990. Personal communication.
Wherry, E.T. 1935. In Walcott, M.V. Illustrations of North American Pitcher Plants. Smithsonian Institution, Washington, D.C.